Forschung und Entwicklung
Ein Ort stetiger Veränderung
Willkommen in unserer Forschungsabteilung!
Hier halten wir dich über unsere aktuellen Projekte auf dem Laufenden, erklären dir den Hintergrund unserer Technologie bis ins Detail und lassen dich an unserer aktuellen Forschung teilhaben.
Unsere Themen:
Unser seedalive-Keimfähigkeitsschnelltest im Detail
Wie gut keimt mein Saatgut? Pflanzensamen kann man ihren Gesundheitszustand nicht ansehen – doch genau dieser ist entscheidend für die Lebensmittelproduktion, Saatgutvermehrung und Züchtung.
Was traditionelle Tests in 7-14 Tagen ermitteln, prognostiziert seedalive verlässlich in nur 3-4 Stunden!
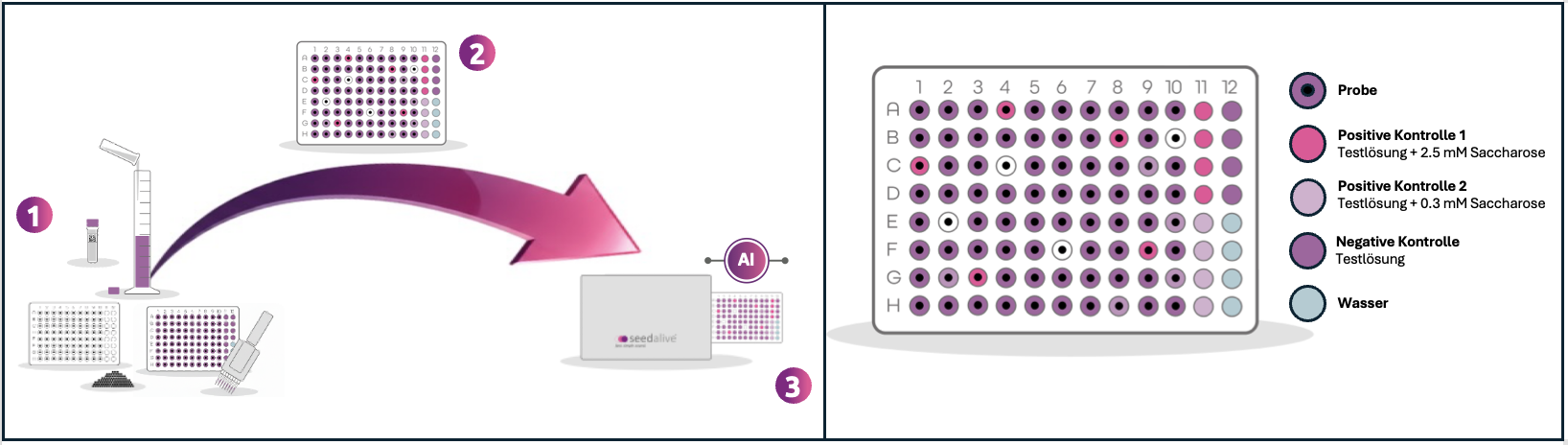
Der seedalive-Bioassay in drei einfachen Schritten:
- Inkubation – Einzelne Samen werden mit dem Seed Dispenser in Mikrotiterplatten vorgelegt, die Testlösung mit einer Mehrkanalpipette appliziert und die Proben 3 – 4 Stunden bei 21°C inkubiert. Die seedalive-Lösung setzt sich aus einem Redox-Indikator (Resazurin) und einem Mikroorganismus (Hefe) zusammen. Zur gleichmäßigen Benetzung der Samen enthält die Lösung außerdem ein Detergens (Superspreiter).
- Reaktion – Je nach Gesundheitszustand der einzelnen Samen werden unterschiedliche Mengen an sameneigenen Substanzen ausgewaschen. Die Mikroorganismen verstoffwechseln diese Verbindungen und verursachen dadurch den Farbumschlag. Zur Sicherstellung der biologischen Funktionalität des Bioassays befinden sich auf jeder Testplatte Negativ- und Positivkontrollen. Negativkontrollen prüfen die ausbleibende Farbveränderung ohne ein Substrat, Positivkontrollen prüfen die erwartete Farbveränderung durch zwei Konzentrationen eines definierten Substrats (Saccharose).
- KI-Analyse – Unsere trainierten KI-Modelle analysieren die photometrisch gemessenen Farbveränderungen und generieren präzise Keimungsvorhersagen. Diese Korrelationszusammenhänge zwischen Farbveränderung und Keimungserfolg wurden mittels umfangreicher Datensätze von zehntausenden von Samen (unterschiedlicher Qualität) pro Kultur trainiert und validiert.
Hintergrund
Was passiert im seedalive-Test?
Trockene Samen befinden sich im Ruhezustand – ihre Zellstrukturen sind „eingepackt", nicht funktionsfähig, aber perfekt lagerfähig. Der entscheidende Moment: beim ersten Wasserkontakt werden die Zellstrukturen reaktiviert.
Besonders kritisch sind dabei die Membransysteme – sie umschließen jede Zelle und grenzen Reaktionsräume voneinander ab. Diese Membranen müssen ihre selektiv-permeable Eigenschaft wiedererlangen, um ordnungsgemäß zu funktionieren und somit u.a. das Zellinnere von der Umwelt abzugrenzen.
Während der Membranrekonstitution werden neben anorganischen auch organische Substanzen wie Zucker, Proteine und Aminosäuren aus den Zellen ausgewaschen (Eluat). Die Regel: Je älter und geschädigter ein Same, desto langsamer und fehleranfälliger läuft die Rekonstitution ab – und desto mehr Substanzen werden freigesetzt.
Die Mikroorganismen in unserer Testlösung nehmen diese ausgewaschenen Substanzen auf und verstoffwechseln sie. Ihre katabole Stoffwechselaktivität reduziert (über NADH als Reduktionsäquivalent) den Redox-Indikator chemisch: die Lösung wechselt von blau (Resazurin) über pink (Resorufin) zu farblos (Dihydroresorufin). So spiegelt die Lösungsfarbe die Samengesundheit wider und ermöglicht präzise Keimfähigkeitsprognosen.
Die zugrundeliegenden zellulären Prinzipien gelten für alle Pflanzensamen. Morphologische Unterschiede zwischen verschiedenen Kulturen berücksichtigen wir durch artspezifische Protokolle und maßgeschneiderte KI-Modelle
Weiterführende Literatur:
Bhattacharya S, Mummenhoff K (2024). Effective seedbank management to ensure food security and preserve biodiversity. Plant Systematics and Evolution 310: 15. https://doi.org/10.1007/s00606-024-01897-z
Mohammed S, Bhattacharya S, Mummenhoff K (2019). Dead or alive: simple, non-destructive, and predictive monitoring of seedbanks. Trends in Plant Science 24: 783-784. https://www.cell.com/action/showPdf?pii=S1360-1385%2819%2930144-X

Weiterentwicklung des seedalive-Tests:
ein Verfahren, viele Einblicke
Der seedalive-Bioassay ermöglicht eine schnelle und zuverlässige Prognose der relevanten Keimungsparameter (Wurzeldurchbruch, normale Keimung und Keimungsindex – nach ISTA sowie Keimenergie und Wasserempfindlichkeit - nach MEBAK / EBC). Aus Forschungs- und Entwicklungsergebnissen unserer Firma ist jedoch immer klarer geworden, dass auch hier noch nicht die Grenze des Möglichen erreicht ist: mithilfe spezifischer und umfangreicher Trainingsdaten können zusätzlich weitere Zusammenhänge mit unserem Bioassay detektiert werden! Somit arbeiten wir daran, alle relevanten Qualitätseigenschaften des Saatguts zu ermitteln – und das alles in nur einem Test!
.
Nutzungsrechte:
Die im Rahmen des Auftrags erstellten Bilder dürfen vom Kunden zeitlich unbeschränkt für alle eigenen Zwecke frei genutzt und auch an Dritte weitergegeben werden. Eine ausschließliche Nutzung durch den Kunden wird nicht eingeräumt. Alle weiteren Rechte verbleiben beim Urheber.
Geltungsbereich:
Die Nutzung ist geografisch auf Europa beschränkt.
Nutzungsdauer:
Die Nutzungsrechte werden zeitlich unbeschränkt eingeräumt.
Nutzungsarten:
Bilder dürfen für alle Zwecke des Kunden verwendet werden, einschließlich, aber nicht beschränkt auf:
• Printmedien (z. B. Broschüren, Flyer, Plakate)
• Digitale Medien (z. B. Websites, Social Media, Newsletter)
• Präsentationen, interne Kommunikation und Werbung
Foto: David Ebener / fototronik.de")
Fusarium-Infektionen
Fusarium – eine Gattung Toxin-bildender Schlauchpilze – zählt zu den gefährlichsten Phytopathogenen weltweit. Diese Pilze befallen wichtige Kulturen wie Weizen, Hafer, Gerste, Roggen, Tomate, Mais und Banane und beeinträchtigen die Pflanzenentwicklung und verursachen drastische Ertragseinbußen und kontaminierte Nahrungsmittel. Der globale Schaden beläuft sich auf mehrere Milliarden Euro jährlich.
Im Rahmen eines Innovationsförderprogramms (IFP) der NBank Niedersachsen untersuchen wir gemeinsam mit der Hochschule Osnabrück zwei zentrale Fragen: 1) Kann der seedalive-Test Fusarium-Infektionen frühzeitig detektieren? 2) Lässt sich der Keimungserfolg infizierter Samen präzise prognostizieren? Die ersten Ergebnisse sind vielversprechend– beide Ziele scheinen mit der seedalive-Technologie erreichbar zu sein. So konnten wir bereits zeigen, dass die Farbveränderungen im seedalive-Test mit der Intensität der Infektion (gemessen über das Mykotoxin Deoxynivalenol) und dem Keimungserfolg der Samen hoch korrelieren.
Neben den landwirtschaftlichen Herausforderungen, die diese Pilze verursachen, ist das sogenannte „Gushing“-Phänomen im Brauwesen bekannt und wird auf Fusarium-Kontaminationen zurückgeführt. Hierbei kommt es zu spontanem Überschäumen des fertigen Bieres sowie einem unerwünschten Geruch. In diesem Zusammenhang braucht es eine sensitive Methode, um auch kleinste Fusarium-Mengen zu detektieren.
Unsere Durchbrüche in der Früherkennung von Fusarium-Infektionen in Braugerste könnten die Brau- und Landwirtschaft revolutionieren und helfen, Ernteverluste zu minimieren sowie die Lebensmittelsicherheit zu erhöhen.
Vorzeitiger Austrieb
Vorzeitiger Austrieb ist ein wachsendes Problem in der Getreideproduktion. Bei feuchter Witterung keimen Getreidekörner bereits auf der Mutterpflanze. Ein unerwünschter Nebeneffekt jahrzehntelanger Züchtung: die natürliche Dormanz wurde so stark reduziert, dass Samen kaum noch über Mechanismen zur Keimungskontrolle verfügen. Beim anschließenden Trocknen für die Lagerung der Samen erleiden vorzeitig gekeimte Körner schwere Schäden oder sterben ab. Dies führt zu erheblichen Verlusten bei der Keimfähigkeit und Triebkraft der Samen. Gleichzeitig verschlechtert sich die stoffliche Zusammensetzung der Körner erheblich, was zu Qualitätseinbußen bei der späteren Verarbeitung führt.
Solche geschädigten Samen lösen eine starke Reaktion im seedalive-Test aus – selbst bei Körnern, die äußerlich völlig intakt erscheinen. Dies ist besonders wertvoll bei minimal ausgeprägtem Auswuchs, der visuell nicht erkennbar ist (verdeckter Auswuchs). Die präzise Detektion und Quantifizierung der Qualitätsminderung ist enorm wichtig – bevor es zu spät ist.
")
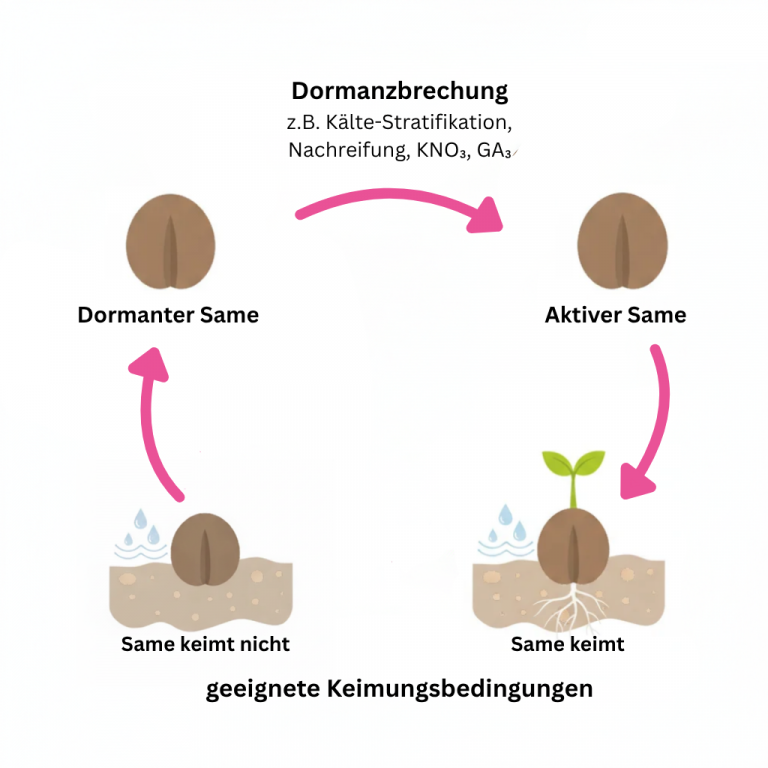
Dormanz-Detektion
Dormanz (Keimruhe) ist ein natürlicher Regulationsmechanismus des Samens, der die Keimung verhindert, obwohl geeignete Bedingungen zum Keimen herrschen. Diverse Regulationsmechanismen blockieren den Keimprozess und stellen sicher, dass die Entwicklung der jungen Pflanze an geeignetem Ort und zur geeigneten Zeit stattfindet. Dies kann unter anderem über die Perzeption eines relevanten Schlüsselreizes realisiert werden: Kälte beispielsweise signalisiert den Winter, die nachfolgende Wärme kündigt dann das Frühjahr als idealen Keimungszeitpunkt an (in gemäßigtem Klima). Oder langanhaltende Regenfälle waschen Hemmstoffe aus der Samenschale und signalisieren die Regenzeit (in sub-tropischem Klima).
Klassische Dormanz-Tests sind zeit- und arbeitsintensiv, und der Dormanz-Grad ist für viele Entscheidungsprozesse in der Saatgutbranche entscheidend. Hier setzt seedalive an: Unsere speziell trainierten KI-Modelle können dormante Samen bereits im seedalive-Test identifizieren. Die vorläufigen Ergebnisse sind vielversprechend und lassen einen echten Durchbruch erwarten – schnelle, zuverlässige Dormanz-Bestimmung statt wochenlanger Standardtests.
Publikationen unter Beteiligung des seedalive-Teams zur Samen- und Keimungsbiologie
Varnskühler J, Wellmann K, Lingen J, Sperber K, Mummenhoff K (2025) Rapid quality assessment of malting barley. Brauwelt International IV: 202 – 205. Fachverlag Hans Carl. https://brauwelt.com/en/topics/raw-materials/648422-rapid-quality-assessment-of-malting-barley
Varnskühler J, Wellmann K, Lingen J, Sperber K, Mummenhoff K. (2025) Schnelle Qualitätskontrolle von Braugerste. Brauwelt17: 606 – 609. Fachverlag Hans Carl. https://brauwelt.com/de/themen/rohstoffe/648390-schnelle-qualit%C3%A4tskontrolle-von-braugerste
Steinbrecher T, Bhattacharya S, Binder J, Kleemeier K, Prezesdzink F, Gröne F, Jacoblinnert K, Mummenhoff K, Leubner-Metzger G (2025) Comparative pericarp biomechanics and germination physiology of Raphanus raphanistrum and Raphanus pugioniformis indehicent fruits. Annals of Botany 135: 977-990. https://doi.org/10.1093/aob/mcaf015
Mohammed S, Steinbrecher T, Leubner-Metzger G, Mummenhoff K (2025) Differential primary seed and fruit dispersal mechanisms and dispersal biomechanics in invasive dehiscent and indehiscent-fruited Lepidium species. Plants 14: 446. https://doi.org/10.3390/plants14030446
Mohammed S, Mummenhoff K (2025) More than just a shell: indehiscent fruits drive drought-tolerant germination in invasive Lepidium species. Plants 14: 1517. https://doi.org/10.3390/plants14101517
Mohammed S, Mummenhoff K (2025) Germination under temperature stress facilitates invasion in indehiscent Lepidium species.Agriculture 15: 1078. https://doi.org/10.3390/agriculture15101078
Mohammed S, Mummenhoff K. (2025) Functional divergence exists in mucilage-mediated seed dispersal, but not in germination of myxospermic Lepidium campestre and Lepidium draba (Brassicaceae). Acta Oecologica 125: 104042. https://doi.org/10.1016/j.actao.2024.104042
Mummenhoff k, Gröne F, Sperber K, Wellmann K, Varnskühler J (2024). Tot oder lebendig? DIY Keimfähigkeitsschnelltest für Pflanzensamen als Kit: Neue Anwendungsbereiche des seedalive Tests. VDLUFA Schriftenreihe 81, Kongressband.
Bhattacharya S, Mummenhoff K. (2024). Effective seedbank management to ensure food security and preserve biodiversity. Plant Systematics and Evolution 310: 15. https://doi.org/10.1007/s00606-024-01897-z
Chandler JO, Wilhelmsson PKI, Fernandez-Pozo N, Graeber K, Arshad W, Pérez M, Steinbrecher T, Ullrich KK, Nguyen T-P, Mérai Z, Mummenhoff K, Theißen G, Strnad M, Mittelsten Scheid O, Schranz ME Petřík I, Tarkowská D, Novák O, Rensing SA, Leubner-Metzger G. (2024). The dimorphic diaspore model Aethionema arabicum (Brassicaceae): Distinct molecular and morphological control of responses to parental and germination temperatures. The Plant Cell 12. https://pmc.ncbi.nlm.nih.gov/articles/PMC11218780/pdf/koae085.pdf
Wellmann K, Varnskühler J, Leubner-Metzger G, Mummenhoff K. (2023). Maize grain germination is accompanied by acidification of the environment. Agronomy 13: 1819. https://doi.org/10.3390/agronomy13071819
Mohammed S, Bhattacharya S, Gesing MA, Klupsch K, Theißen G, Mummenhoff K, Müller C (2020). Morphologically and physiologically diverse fruits of two Lepidium species differ in allocation of glucosinolates into immature and mature seed and pericarp. PLoSONE 15. https://journals.plos.org/plosone/article/file?id=10.1371/journal.pone.0227528&type=printable
Ziffer-Berger J, Waitz Y, Behar E, Ben Joseph O, Bezalel L, Wasserstrom H, Bajpai P K, Bhattacharya S, Przesdzink F, Westberg E, Mummenhoff K, Barazani O. (2020). Seed dispersal of wild radishes and its association with within-population spatial distribution. BMC Ecology 20: file:///C:/Users/Mummenhoff/Downloads/s12898-020-00297-4-2.pdf
Bhattacharya S, Sperber K, Özüdoğru B, Leubner-Metzger G, Mummenhoff K (2019). In the right place at the right time: Naturally- primed dramatic plasticity in the life strategy of dimorphic Aethionema arabicum (Brassicaceae) facilitates colonizing the optimal habitat. Scientific Reports 9: 16108. https://doi.org/10.1038/s41598-019-52520-y
Mohammed S, Bhattacharya S, Mummenhoff K (2019). Dead or alive: simple, non-destructive, and predictive monitoring of seedbanks. Trends in Plant Science 24: 783-784. https://www.cell.com/action/showPdf?pii=S1360-1385%2819%2930144-X
Mohammed S, Turečková V, Tarkowská D, Strnad M, Mummenhoff K, Leubner-Metzger G. (2019). Pericarp-mediated chemical dormancy controls the fruit germination of the invasive Hoary Cress (Lepidium draba), but not of Hairy Whitetop (Lepidium appelianum). Weed Science 67: 560-571. https://doi.org/10.1017/wsc.2019.33
Arshad W, Sperber K, Steinbrecher T, Nichols B, Jansen VA, Leubner-Metzger G, Mummenhoff K (2018). Dispersal biophysics and adaptive significance of dimorphic diaspores in the annual Aethionema arabicum (Brassicaceae). New Phytologist221: 1434-1446. https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.15490
Bhattacharya S, Mayland-Quellhorst S, Müller C, Mummenhoff K. (2018). Two-tier morpho-chemical defense tactic in Aethionema via fruit-morph plasticity and glucosinolates allocation in diaspores. Plant, Cell & Environment 42: 1381-1392. https://onlinelibrary.wiley.com/doi/epdf/10.1111/pce.13462
Sperber K, Steinbrecher T, Graeber K, Scherer G, Clausing S, Wiegand N, Hourston JE, Kurre R, Leubner-Metzger G, Mummenhoff K (2017). Fruit fracture biomechanics and the release of Lepidium didymum pericarp-imposed mechanical dormancy by fungi. Nature Communications 8: 1868. file:///C:/Users/Mummenhoff/Downloads/s41467-017-02051-9.pdf
Lenser T, et al, Theißen G, Mummenhoff K, Leubner-Metzger G (2016). Developmental control and plasticity of fruit and seed dimorphism in Aethionema arabicum. Plant Physiology 172: 1691–1707. https://doi.org/10.1104/pp.16.00838
Özüdoğru B, Akaydın G, Erik S, Mummenhoff K (2016). Seed morphology of Ricotia L (Brassicaceae) and its phylogenetic and systematic implication. Flora 222: 60–67. https://doi.org/10.1016/j.flora.2016.03.013
Graeber K, Linkies A, Steinbrecher T, Mummenhoff K, Tarkowská D, Turečková V, Ignatz M, Sperber K, Voegele A, de Jong H, Urbanová T, Strnad M, Leubner-Metzger G (2014). DELAY OF GERMINATION 1 mediates a conserved coat dormancy mechanism for the temperature- and gibberellin-dependent control of seed germination. Proceedings of the National Academy of Science of the USA 111: E3571–E3580. https://www.pnas.org/doi/epdf/10.1073/pnas.1403851111
Mühlhausen A, Lenser T, Mummenhoff K, Theißen G (2013). Evidence that an evolutionary transition from dehiscent to indehiscent fruits in Lepidium (Brassicaceae) was caused by a change in the control of valve margin identity genes. Plant Journal75: 824–835. https://doi.org/10.1111/tpj.12079
Graeber K, Büttner-Mainik A, Sperber K, Mummenhoff K, Leubner-Metzger G (2013). Spatio-temporal seed development analysis provide insight into primary dormancy induction and evolution of the Lepidium DELAY OF GERMINATION 1 genes. Plant Physiology 161: 1903-1917. https://pmc.ncbi.nlm.nih.gov/articles/PMC3613464/pdf/1903.pdf
Mummenhoff K, Polster A, Mühlhausen A, Theißen G (2009). Lepidium as a model system for studying the evolution of fruit development in Brassicaceae. Journal of Experimental Botany 60: 1503-1513. https://doi.org/10.1093/jxb/ern304
Mummenhoff K, Theißen G, Mühlhausen A (2008). Tutti Frutti in Lepidium — Comparative analysis of fruit dehiscence/indehiscence in Brassicaceae. South African Journal of Botany 74: 373-374. https://DOI:10.1016/J.SAJB.2008.01.086
Moazzeni H, Zarre S, Al-Shehbaz IA, Mummenhoff K (2007). Seed-coat microsculpturing and its systematic application in Isatis (Brassicaceae) and allied genera in Iran. Flora 202: 447–454. https://doi.org/10.1016/j.flora.2006.10.004