Research and Development
A place of constant change
Welcome to our research department!
Here, we’ll keep you up to date with our current projects, explain the background to our technology in detail, and give you an insight into our latest research.
Our topics:
Our seedalive rapid germination test in detail
How well do my seeds germinate? You can’t tell how healthy plant seeds are just by looking at them – yet this is precisely what is crucial for food production, seed propagation and breeding.
What traditional tests take 7–14 days to determine, seedalive reliably predicts in just 3–4 hours!
The seedalive bioassay in three simple steps:
- Incubation – Individual seeds are dispensed into microtitre plates using the Seed Dispenser, the test solution is applied with a multichannel pipette, and the samples are incubated for 3–4 hours at 21°C. The seedalive solution consists of a redox indicator (resazurin) and a microorganism (yeast). To ensure uniform wetting of the seeds, the solution also contains a detergent (super-spreader).
- Reaction – Depending on the health status of the individual seeds, varying amounts of the seeds’ own substances are leached out. The microorganisms metabolise these compounds, thereby causing the colour change. To ensure the biological functionality of the bioassay, each test plate contains negative and positive controls. Negative controls verify the absence of a colour change in the absence of a substrate, whilst positive controls verify the expected colour change using two concentrations of a defined substrate (sucrose).
- AI analysis – Our trained AI models analyse the photometrically measured colour changes and generate precise germination predictions. These correlations between colour change and germination success were trained and validated using extensive datasets comprising tens of thousands of seeds (of varying quality) per culture.

Background
What happens during the seedalive test?
Dry seeds are in a dormant state – their cellular structures are ‘packaged away’, non-functional but perfectly suited to storage. The crucial moment: upon first contact with water, the cellular structures are reactivated.
The membrane systems are particularly critical in this process – they enclose each cell and separate reaction compartments from one another. These membranes must regain their selectively permeable properties in order to function properly and thus, amongst other things, isolate the cell interior from the environment.
During membrane reconstitution, both inorganic and organic substances such as sugars, proteins and amino acids are washed out of the cells (eluate). The rule is: the older and more damaged a seed is, the slower and more error-prone the reconstitution process is – and the more substances are released.
The microorganisms in our test solution take up these leached substances and metabolise them. Their catabolic metabolic activity chemically reduces the redox indicator (via NADH as a reducing equivalent): the solution changes colour from blue (resazurin) to pink (resorufin) to colourless (dihydroresorufin). In this way, the colour of the solution reflects seed health and enables precise germination predictions.
The underlying cellular principles apply to all plant seeds. We account for morphological differences between different crops through species-specific protocols and bespoke AI models.
Further Literature:
Bhattacharya S, Mummenhoff K (2024). Effective seedbank management to ensure food security and preserve biodiversity. Plant Systematics and Evolution 310: 15. https://doi.org/10.1007/s00606-024-01897-z
Mohammed S, Bhattacharya S, Mummenhoff K (2019). Dead or alive: simple, non-destructive, and predictive monitoring of seedbanks. Trends in Plant Science 24: 783-784. https://www.cell.com/action/showPdf?pii=S1360-1385%2819%2930144-X

Further development of the seedalive test:
One method, many insights
The seedalive bioassay enables a rapid and reliable prediction of the relevant germination parameters (root emergence, normal germination and germination index – according to ISTA – as well as germination energy and water sensitivity – according to MEBAK / EBC). However, research and development findings from our company have made it increasingly clear that we have not yet reached the limits of what is possible in this area either: with the help of specific and comprehensive training data, further correlations can be detected using our bioassay! We are therefore working to determine all the relevant quality characteristics of the seed – all in a single test!
.
Nutzungsrechte:
Die im Rahmen des Auftrags erstellten Bilder dürfen vom Kunden zeitlich unbeschränkt für alle eigenen Zwecke frei genutzt und auch an Dritte weitergegeben werden. Eine ausschließliche Nutzung durch den Kunden wird nicht eingeräumt. Alle weiteren Rechte verbleiben beim Urheber.
Geltungsbereich:
Die Nutzung ist geografisch auf Europa beschränkt.
Nutzungsdauer:
Die Nutzungsrechte werden zeitlich unbeschränkt eingeräumt.
Nutzungsarten:
Bilder dürfen für alle Zwecke des Kunden verwendet werden, einschließlich, aber nicht beschränkt auf:
• Printmedien (z. B. Broschüren, Flyer, Plakate)
• Digitale Medien (z. B. Websites, Social Media, Newsletter)
• Präsentationen, interne Kommunikation und Werbung
Foto: David Ebener / fototronik.de")
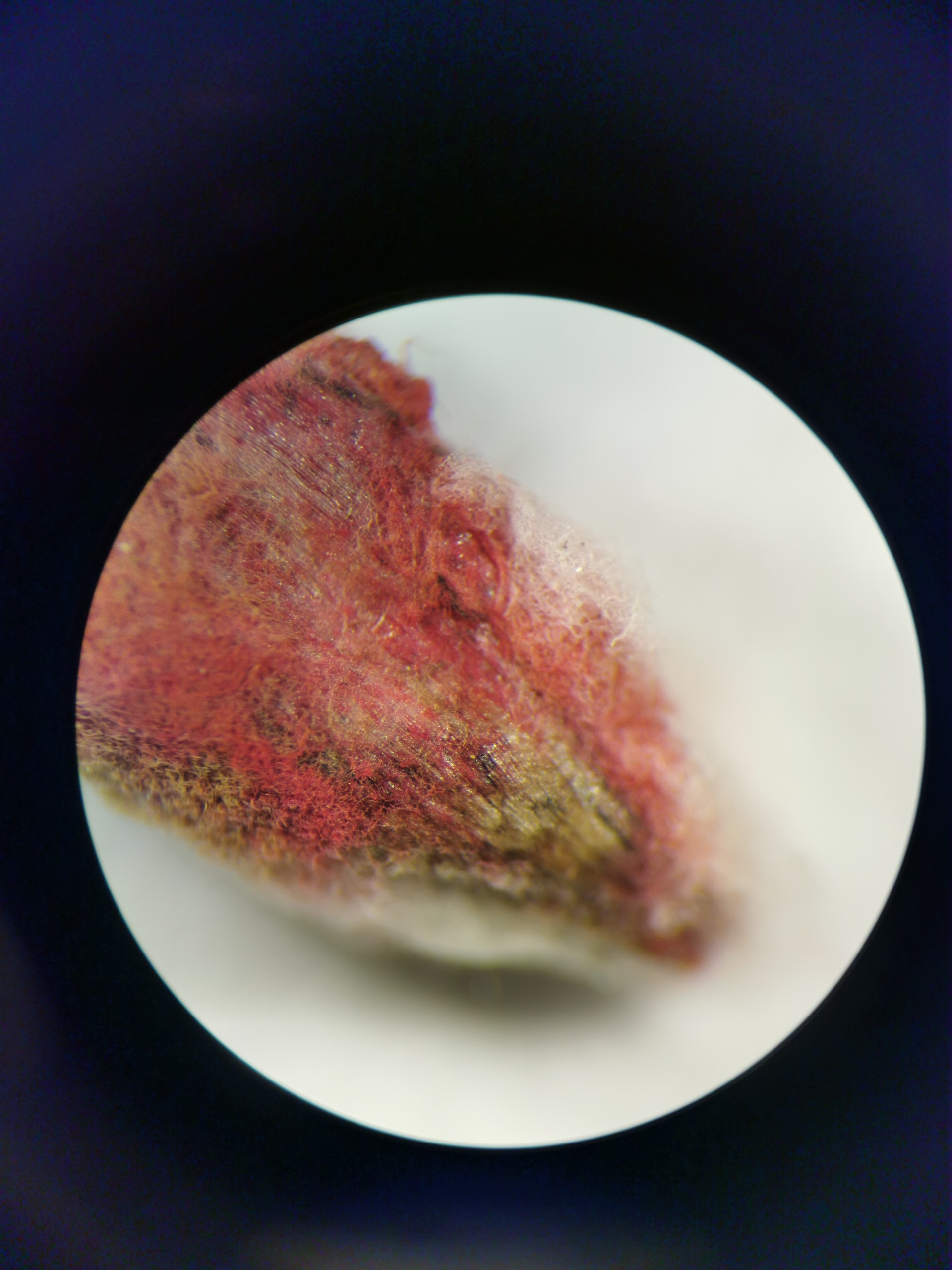
Fusarium infections
Fusarium – a genus of toxin-producing fungi – is one of the most dangerous plant pathogens in the world. These fungi infect key crops such as wheat, oats, barley, rye, tomatoes, maize and bananas, impairing plant development and causing drastic yield losses and food contamination. The global damage amounts to several billion euros annually.
As part of an innovation funding programme (IFP) run by NBank Niedersachsen, we are collaborating with Osnabrück University of Applied Sciences to investigate two key questions: 1) Can the seedalive test detect Fusarium infections at an early stage? 2) Can the germination success of infected seeds be accurately predicted? The initial results are promising – both objectives appear to be achievable with seedalive technology. We have already been able to demonstrate that the colour changes observed in the seedalive test correlate strongly with the intensity of the infection (measured via the mycotoxin deoxynivalenol) and the germination success of the seeds.
In addition to the agricultural challenges caused by these fungi, the so-called ‘gushing’ phenomenon is well known in the brewing industry and is attributed to Fusarium contamination. This results in spontaneous foaming of the finished beer as well as an undesirable odour. In this context, a sensitive method is required to detect even the smallest amounts of Fusarium.
Our breakthroughs in the early detection of Fusarium infections in malting barley could revolutionise the brewing and agricultural sectors, helping to minimise crop losses and improve food safety.
Preharvest sprouting
Premature sprouting is a growing problem in cereal production. In damp weather, cereal grains germinate whilst still on the parent plant. An undesirable side-effect of decades of breeding: natural dormancy has been reduced to such an extent that seeds now have virtually no mechanisms left to control germination. During the subsequent drying process for seed storage, prematurely germinated grains suffer severe damage or die off. This leads to significant losses in the seeds’ germination capacity and vigour. At the same time, the chemical composition of the grains deteriorates considerably, resulting in a loss of quality during subsequent processing.
Such damaged seeds trigger a strong reaction in the seedalive test – even in grains that appear completely intact on the outside. This is particularly valuable in cases of minimal sprouting that is not visually detectable (hidden sprouting). The precise detection and quantification of this reduction in quality is of paramount importance – before it is too late.
")

Dormancy detection
Dormancy is a natural regulatory mechanism in seeds that prevents germination even when suitable conditions for germination are present. Various regulatory mechanisms block the germination process and ensure that the young plant develops in the right place and at the right time. This can be achieved, amongst other things, through the perception of a relevant key stimulus: cold, for example, signals winter, whilst the subsequent warmth then heralds spring as the ideal time for germination (in temperate climates). Or prolonged rainfall washes inhibitors out of the seed coat and signals the rainy season (in subtropical climates).
Traditional dormancy tests are time-consuming and labour-intensive, and the degree of dormancy is crucial for many decision-making processes in the seed industry. This is where seedalive comes in: our specially trained AI models can identify dormant seeds as early as the seedalive test. The preliminary results are promising and suggest a genuine breakthrough is on the horizon – fast, reliable dormancy determination instead of weeks of standard tests.
Publications on seed and germination biology involving the seedalive team
Varnskühler J, Wellmann K, Lingen J, Sperber K, Mummenhoff K (2025) Rapid quality assessment of malting barley. Brauwelt International IV: 202 – 205. Fachverlag Hans Carl. https://brauwelt.com/en/topics/raw-materials/648422-rapid-quality-assessment-of-malting-barley
Varnskühler J, Wellmann K, Lingen J, Sperber K, Mummenhoff K. (2025) Schnelle Qualitätskontrolle von Braugerste. Brauwelt17: 606 – 609. Fachverlag Hans Carl. https://brauwelt.com/de/themen/rohstoffe/648390-schnelle-qualit%C3%A4tskontrolle-von-braugerste
Steinbrecher T, Bhattacharya S, Binder J, Kleemeier K, Prezesdzink F, Gröne F, Jacoblinnert K, Mummenhoff K, Leubner-Metzger G (2025) Comparative pericarp biomechanics and germination physiology of Raphanus raphanistrum and Raphanus pugioniformis indehicent fruits. Annals of Botany 135: 977-990. https://doi.org/10.1093/aob/mcaf015
Mohammed S, Steinbrecher T, Leubner-Metzger G, Mummenhoff K (2025) Differential primary seed and fruit dispersal mechanisms and dispersal biomechanics in invasive dehiscent and indehiscent-fruited Lepidium species. Plants 14: 446. https://doi.org/10.3390/plants14030446
Mohammed S, Mummenhoff K (2025) More than just a shell: indehiscent fruits drive drought-tolerant germination in invasive Lepidium species. Plants 14: 1517. https://doi.org/10.3390/plants14101517
Mohammed S, Mummenhoff K (2025) Germination under temperature stress facilitates invasion in indehiscent Lepidium species.Agriculture 15: 1078. https://doi.org/10.3390/agriculture15101078
Mohammed S, Mummenhoff K. (2025) Functional divergence exists in mucilage-mediated seed dispersal, but not in germination of myxospermic Lepidium campestre and Lepidium draba (Brassicaceae). Acta Oecologica 125: 104042. https://doi.org/10.1016/j.actao.2024.104042
Mummenhoff k, Gröne F, Sperber K, Wellmann K, Varnskühler J (2024). Tot oder lebendig? DIY Keimfähigkeitsschnelltest für Pflanzensamen als Kit: Neue Anwendungsbereiche des seedalive Tests. VDLUFA Schriftenreihe 81, Kongressband.
Bhattacharya S, Mummenhoff K. (2024). Effective seedbank management to ensure food security and preserve biodiversity. Plant Systematics and Evolution 310: 15. https://doi.org/10.1007/s00606-024-01897-z
Chandler JO, Wilhelmsson PKI, Fernandez-Pozo N, Graeber K, Arshad W, Pérez M, Steinbrecher T, Ullrich KK, Nguyen T-P, Mérai Z, Mummenhoff K, Theißen G, Strnad M, Mittelsten Scheid O, Schranz ME Petřík I, Tarkowská D, Novák O, Rensing SA, Leubner-Metzger G. (2024). The dimorphic diaspore model Aethionema arabicum (Brassicaceae): Distinct molecular and morphological control of responses to parental and germination temperatures. The Plant Cell 12. https://pmc.ncbi.nlm.nih.gov/articles/PMC11218780/pdf/koae085.pdf
Wellmann K, Varnskühler J, Leubner-Metzger G, Mummenhoff K. (2023). Maize grain germination is accompanied by acidification of the environment. Agronomy 13: 1819. https://doi.org/10.3390/agronomy13071819
Mohammed S, Bhattacharya S, Gesing MA, Klupsch K, Theißen G, Mummenhoff K, Müller C (2020). Morphologically and physiologically diverse fruits of two Lepidium species differ in allocation of glucosinolates into immature and mature seed and pericarp. PLoSONE 15. https://journals.plos.org/plosone/article/file?id=10.1371/journal.pone.0227528&type=printable
Ziffer-Berger J, Waitz Y, Behar E, Ben Joseph O, Bezalel L, Wasserstrom H, Bajpai P K, Bhattacharya S, Przesdzink F, Westberg E, Mummenhoff K, Barazani O. (2020). Seed dispersal of wild radishes and its association with within-population spatial distribution. BMC Ecology 20: file:///C:/Users/Mummenhoff/Downloads/s12898-020-00297-4-2.pdf
Bhattacharya S, Sperber K, Özüdoğru B, Leubner-Metzger G, Mummenhoff K (2019). In the right place at the right time: Naturally- primed dramatic plasticity in the life strategy of dimorphic Aethionema arabicum (Brassicaceae) facilitates colonizing the optimal habitat. Scientific Reports 9: 16108. https://doi.org/10.1038/s41598-019-52520-y
Mohammed S, Bhattacharya S, Mummenhoff K (2019). Dead or alive: simple, non-destructive, and predictive monitoring of seedbanks. Trends in Plant Science 24: 783-784. https://www.cell.com/action/showPdf?pii=S1360-1385%2819%2930144-X
Mohammed S, Turečková V, Tarkowská D, Strnad M, Mummenhoff K, Leubner-Metzger G. (2019). Pericarp-mediated chemical dormancy controls the fruit germination of the invasive Hoary Cress (Lepidium draba), but not of Hairy Whitetop (Lepidium appelianum). Weed Science 67: 560-571. https://doi.org/10.1017/wsc.2019.33
Arshad W, Sperber K, Steinbrecher T, Nichols B, Jansen VA, Leubner-Metzger G, Mummenhoff K (2018). Dispersal biophysics and adaptive significance of dimorphic diaspores in the annual Aethionema arabicum (Brassicaceae). New Phytologist221: 1434-1446. https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.15490
Bhattacharya S, Mayland-Quellhorst S, Müller C, Mummenhoff K. (2018). Two-tier morpho-chemical defense tactic in Aethionema via fruit-morph plasticity and glucosinolates allocation in diaspores. Plant, Cell & Environment 42: 1381-1392. https://onlinelibrary.wiley.com/doi/epdf/10.1111/pce.13462
Sperber K, Steinbrecher T, Graeber K, Scherer G, Clausing S, Wiegand N, Hourston JE, Kurre R, Leubner-Metzger G, Mummenhoff K (2017). Fruit fracture biomechanics and the release of Lepidium didymum pericarp-imposed mechanical dormancy by fungi. Nature Communications 8: 1868. file:///C:/Users/Mummenhoff/Downloads/s41467-017-02051-9.pdf
Lenser T, et al, Theißen G, Mummenhoff K, Leubner-Metzger G (2016). Developmental control and plasticity of fruit and seed dimorphism in Aethionema arabicum. Plant Physiology 172: 1691–1707. https://doi.org/10.1104/pp.16.00838
Özüdoğru B, Akaydın G, Erik S, Mummenhoff K (2016). Seed morphology of Ricotia L (Brassicaceae) and its phylogenetic and systematic implication. Flora 222: 60–67. https://doi.org/10.1016/j.flora.2016.03.013
Graeber K, Linkies A, Steinbrecher T, Mummenhoff K, Tarkowská D, Turečková V, Ignatz M, Sperber K, Voegele A, de Jong H, Urbanová T, Strnad M, Leubner-Metzger G (2014). DELAY OF GERMINATION 1 mediates a conserved coat dormancy mechanism for the temperature- and gibberellin-dependent control of seed germination. Proceedings of the National Academy of Science of the USA 111: E3571–E3580. https://www.pnas.org/doi/epdf/10.1073/pnas.1403851111
Mühlhausen A, Lenser T, Mummenhoff K, Theißen G (2013). Evidence that an evolutionary transition from dehiscent to indehiscent fruits in Lepidium (Brassicaceae) was caused by a change in the control of valve margin identity genes. Plant Journal75: 824–835. https://doi.org/10.1111/tpj.12079
Graeber K, Büttner-Mainik A, Sperber K, Mummenhoff K, Leubner-Metzger G (2013). Spatio-temporal seed development analysis provide insight into primary dormancy induction and evolution of the Lepidium DELAY OF GERMINATION 1 genes. Plant Physiology 161: 1903-1917. https://pmc.ncbi.nlm.nih.gov/articles/PMC3613464/pdf/1903.pdf
Mummenhoff K, Polster A, Mühlhausen A, Theißen G (2009). Lepidium as a model system for studying the evolution of fruit development in Brassicaceae. Journal of Experimental Botany 60: 1503-1513. https://doi.org/10.1093/jxb/ern304
Mummenhoff K, Theißen G, Mühlhausen A (2008). Tutti Frutti in Lepidium — Comparative analysis of fruit dehiscence/indehiscence in Brassicaceae. South African Journal of Botany 74: 373-374. https://DOI:10.1016/J.SAJB.2008.01.086
Moazzeni H, Zarre S, Al-Shehbaz IA, Mummenhoff K (2007). Seed-coat microsculpturing and its systematic application in Isatis (Brassicaceae) and allied genera in Iran. Flora 202: 447–454. https://doi.org/10.1016/j.flora.2006.10.004